
エアコンが感染リスクを上げる理由|夏の体調不良と風邪の意外な関係


前回は、シアル酸およびインフルエンザウイルス感染機構がどのように発見されたのかについてご紹介しました。シアル酸が宿主細胞の受容体として機能し、インフルエンザウイルスがこれを利用して感染することが明らかになったことで、ウイルス感染の仕組みが大きく解明されました。
今回は、その感染を担うヘマグルチニン(Hemagglutinin:HA)とノイラミニダーゼ(Neuraminidase:NA)という2つの重要な糖タンパク質について解説します。これらはインフルエンザウイルス表面に存在するスパイク状の構造であり、それぞれ異なる役割を担いながら、感染の成立からウイルスの拡散までを支えています。また、HAやNAの変異は、季節性インフルエンザの流行やパンデミックの発生とも深く関わっています。
ヘマグルチニン(HA)は、インフルエンザウイルスが宿主細胞へ感染する最初の段階である細胞への吸着と、その後のエンドサイトーシス(細胞がウイルスを取り込む仕組み)を経た膜融合に関与する膜糖タンパク質です。そのため、インフルエンザウイルスの感染性や宿主特異性を決定する最も重要な分子の一つと考えられています。
同時に、HAは宿主の免疫系に最も認識されやすい抗原でもあり、現在のインフルエンザワクチン開発における主要な標的となっています。HA分子には、受容体結合部位、膜融合ペプチド、抗原決定部位などが存在し、これらの部位に生じる変異は感染性や流行性に大きな影響を与えます。実際、この約100年間に発生した主なパンデミックの多くは、新たなHAサブタイプの出現と密接に関係していることが分かっています。
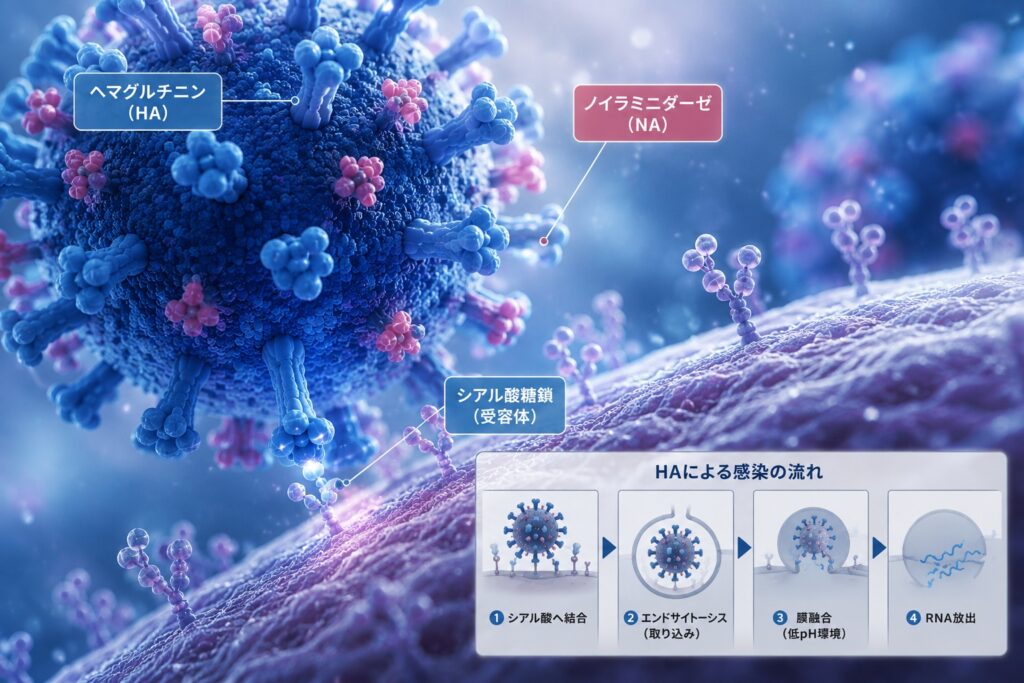
HAは、三量体(トリマー)を形成する糖タンパク質であり、約550個のアミノ酸からなるHA0という前駆体として合成されます。その後、宿主のプロテアーゼによってHA1とHA2に切断されることで、初めて感染能力を持つようになります。この切断が起こらなければ、ウイルスは感染することができません。
HAは頭部(head)と茎部(stem)に大きく分けられます。HA1は主に頭部を構成し、シアル酸糖鎖と結合する受容体結合部位を担っています。一方、HA2はウイルス膜の近くに位置し、膜融合ペプチドを含んでいます。
通常、HAは中性の環境では安定した構造を保っています。しかし、ウイルスが細胞内へ取り込まれた後、エンドソーム内が酸性(低pH)になるとHAは大きく構造を変化させます。この低pH依存的な構造変化によってウイルス膜と細胞膜が融合し、ウイルスRNAが細胞内へ放出されます。この一連の過程は、インフルエンザウイルスの感染成立における中心的な仕組みです。
HAの最も重要な働きの一つが、宿主細胞表面のシアル酸含有糖鎖への結合です。
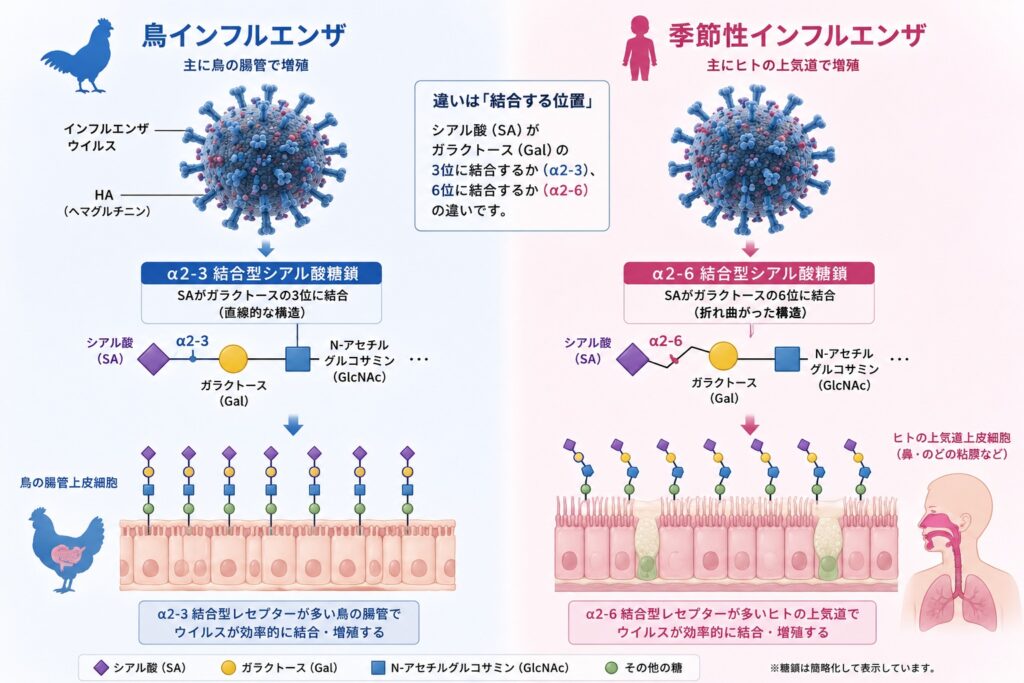
シアル酸とガラクトースの結合様式には、α2-3結合(鳥型)とα2-6結合(ヒト型)があります。鳥類の腸管には主にα2-3型、ヒトの上気道には主にα2-6型が分布しており、この違いがウイルスの宿主特異性を決定しています。
HAの受容体結合部位は浅いポケット状の構造をしており、わずかなアミノ酸の変異でも結合するシアル酸の種類が大きく変化することがあります。このため、鳥インフルエンザウイルスがヒト型(α2-6)のシアル酸へ結合できる能力を獲得すると、人への感染や人から人への感染が起こる可能性が高まり、新たなパンデミックにつながる危険性があります。現在も世界中でインフルエンザウイルスの変異が継続的に監視されているのは、このためです。
HAは宿主細胞への結合を担うだけでなく、宿主免疫系が最も認識しやすい主要抗原でもあります。その頭部に存在する抗原決定部位は中和抗体の標的となりますが、この領域は変異しやすいという特徴があります。そのため、ウイルスは免疫から逃れながら流行を繰り返しています。
HAの抗原変異には、大きく「抗原ドリフト」と「抗原シフト」の2種類があります。
抗原ドリフトは、遺伝子の点突然変異が少しずつ蓄積することで起こる変化です。この小さな変異が積み重なることで、過去の感染やワクチンによって獲得した免疫が十分に働かなくなり、季節性インフルエンザが毎年流行する原因となっています。
一方、抗原シフトは、異なるインフルエンザウイルス同士が遺伝子を交換し、新しいHAサブタイプを持つウイルスが出現する現象です。人類が免疫を持たない新しいHAを持つため、世界的な大流行(パンデミック)を引き起こす可能性があります。
実際に、1918年のスペインインフルエンザ(H1N1)、1957年のアジア風邪(H2N2)、1968年の香港風邪(H3N2)、そして2009年の新型インフルエンザ(H1N1)は、いずれも新しいHAサブタイプの出現と密接に関係していました。特に1918年のスペインインフルエンザでは、世界で数千万人規模の死亡者が報告されており、その影響の大きさが知られています。
パンデミックが成立するためには、
という3つの条件が必要と考えられています。水鳥はH1〜H16まで多様なHA亜型を保有していますが、その多くは鳥型レセプター(α2-3結合シアル酸)を認識します。その中で、ヒト型レセプターへの適応を獲得したH1、H2、H3のみがヒト社会に定着してきました。
このように、抗原性は変化しても、シアル酸を認識するという基本的な仕組みは維持されることが、インフルエンザウイルスの大きな特徴です。
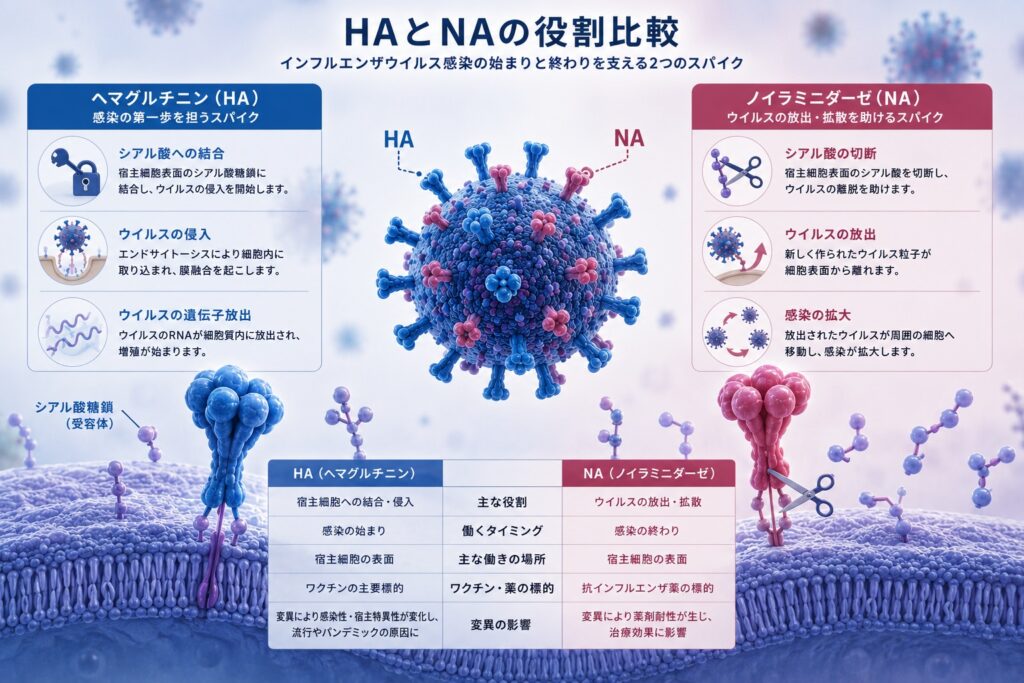
一方、ノイラミニダーゼ(NA)は、シアル酸を切断するシアリダーゼという酵素です。主な役割は、新しく作られたウイルス粒子を感染した細胞から放出し、次の細胞へ感染できるようにすることです。
NAは、膜貫通領域、stalk(柄)領域、そして酵素活性を持つ頭部から構成される四量体タンパク質です。頭部は6枚のβシートからなるプロペラ状の立体構造を形成し、その中心にシアル酸を切断する活性部位があります。
感染した細胞内で増殖したウイルスは、細胞表面のシアル酸糖鎖に結合したままでは周囲へ拡散できません。NAがシアル酸を切断することで初めて、新しく作られたウイルスは細胞から放出され、次の細胞へ感染することができます。このため、NAはウイルス放出に不可欠な酵素として知られています。
さらに近年では、NAは気道粘液中のシアル酸を切断してウイルスの移動を助けることや、細胞内での増殖効率や免疫応答にも関与するなど、多機能なタンパク質であることが明らかになっています。
現在使用されているオセルタミビル(タミフル)、ザナミビル(リレンザ)、ペラミビル(ラピアクタ)などの抗インフルエンザ薬は、シアル酸の遷移状態を模倣して設計されたノイラミニダーゼ阻害薬です。これらはNAの活性部位に結合することでシアル酸の切断を阻害し、新しく作られたウイルスが細胞から放出されるのを防ぎます。
また、HAとNAはそれぞれ独立して働くのではなく、互いの機能が適切なバランスを保つことで、ウイルスは効率よく感染・伝播しています。このバランスが崩れると、ウイルスは十分に増殖・拡散することができません。
近年では、シアル酸糖鎖を模倣した疑似受容体によってウイルスをあらかじめ捕捉する方法や、変異しにくいHA茎部を標的としたユニバーサルワクチンの開発も進められています。これらは、毎年の変異に左右されにくい新しい感染対策として期待されています。
このように、HAとNAはそれぞれ異なる役割を担いながらも、相互に機能的なバランスを保つことでインフルエンザウイルスの感染と伝播を支えています。これらの分子の構造や機能、そして変異の仕組みを理解することは、ワクチンや抗ウイルス薬の開発だけでなく、将来のパンデミック対策を考えるうえでも極めて重要です。今後もHAとNAに関する研究は、感染症対策を支える重要な基盤として発展していくことが期待されています。
静岡県立大学名誉教授・鈴木康夫



